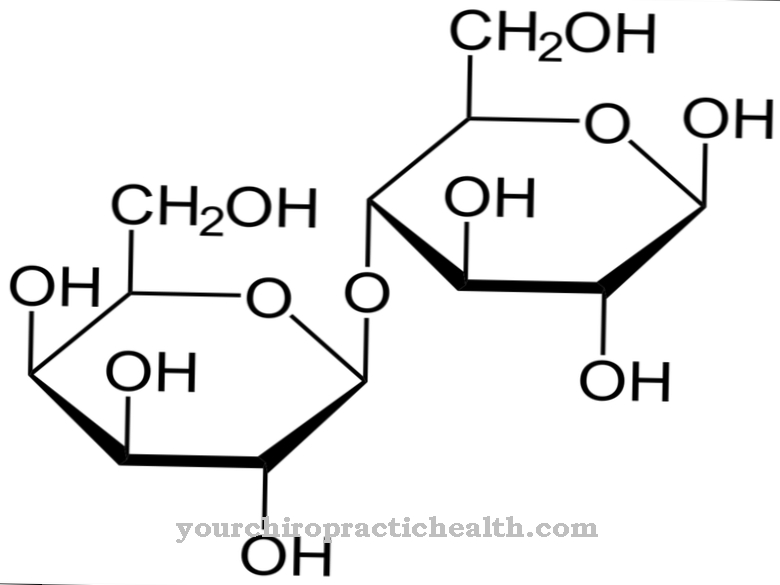
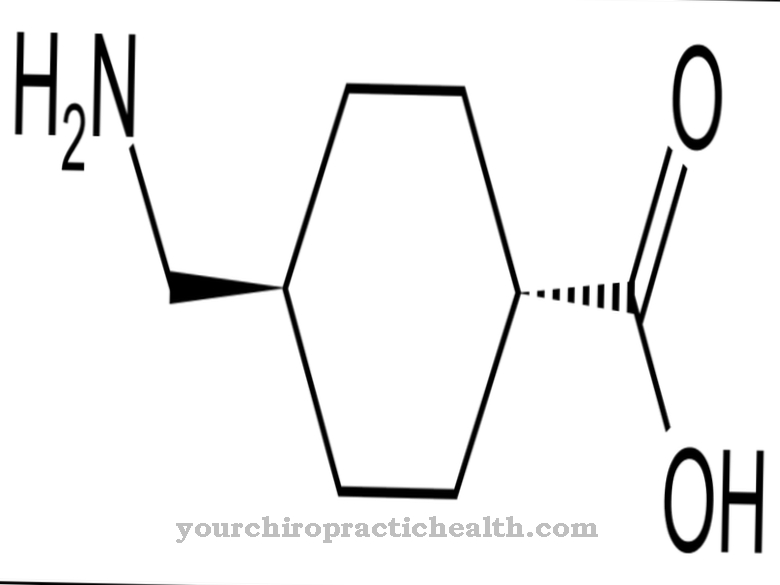
Tsütidiin kuulub nukleosiididesse ja koosneb nukleiinalusest tsütosiinist ja suhkru riboosist. See moodustab vesiniksidemete kaudu guanosiiniga aluspaari. Samuti mängib see keskset rolli pürimidiini metabolismis.
Mis on tsütidiin?
Tsütidiin on nukleosiid, mis koosneb tsütosiinist ja riboosist. Lisaks adeniinile, guaniinile ja tümiinile osaleb nukleiinhapete sünteesis ka lämmastikaluseline tsütosiin. Tsütidiini fosforüülimisel saadakse tsütidiinmonofosfaat (CMP), tsütidiindifosfaat (CDP) või tsütidiintrifosfaat (CTP).
Tsütidiinmonofosfaat on RNA nukleotiid. Nukleiinhapete struktuuris osalevad kaks puriini ja kaks pürimidiinalust, kusjuures tümiin vahetatakse RNA-s uratsiili vastu. Adeniin ja guaniin kuuluvad puriini aluste hulka, tümiin, tsütosiin ja uratsiil kuuluvad pürimidiini aluste hulka. Tsütidiindeaminaas võib demineerida tsütidiini uridiiniks. Uridiin on nukleosiid, mis on valmistatud riboosist ja uratsiilist. Seda saab fosforüülida ka uridiinmonofosfaadiks.
Uridiinmonofosfaat on samuti RNA oluline nukleotiid. Lisaks on CDP ja CTP aktiveerivad rühmad ka letsitiini, tsefaliini ja kardiolipiini sünteesil. Puhas tsütidiin on vees lahustuva tahke ainena, mis laguneb temperatuuril 201–220. Pürimidiini nukleosidaasi abil saab seda katalüütiliselt lagundada tsütosiiniks ja riboosiks.
Funktsioon, mõju ja ülesanded
Tsütidiinil on keskne roll pürimidiini metabolismis. Pürimidiin tagab nukleiinhapetes esinevate tsütosiini, tümiini ja uratsiili pürimidiinaluste põhistruktuuri. RNA-s sisalduv tümiin vahetatakse uratsiili vastu.
Uratsiili toodetakse ka tsütidiini deamineerimisel tsütidiindeaminaasiga. Kolme pürimidiinaluse vahelised keemilised muundamised on DNA paranemisprotsesside ja epigeneetiliste muutuste jaoks keskse tähtsusega. Epigeneetika kontekstis muudavad keskkonnamõjud mitmesuguseid omadusi. Geneetiline materjal aga ei muutu. Organismi modifikatsioonimuutused on põhjustatud geenide erinevast ekspressioonist. Keharakkude diferentseerumisprotsessid erinevate rakuliinide ja elundite moodustamiseks esindavad ka epigeneetilist protsessi, sõltuvalt rakutüübist aktiveeritakse või deaktiveeritakse erinevad geenid.
See toimub tsütidiini aluste metüleerimise kaudu DNA-s. Metüleerimise käigus moodustub metüültsütosiin, mida saab deaminatsiooni teel muundada tümiiniks. Komplementaarsel nukleobaas-guaniinil on vastassuunalises topelt ahelas vea äratundmine ja tümiini vahetamine taas tsütosiini vastu. Siiski võib guaniini vahetada ka adeniini vastu, mis põhjustab punktmutatsiooni. Kui metüleerimata tsütosiin deamineeritakse, toodetakse uratsiili. Kuna uratsiil ei ilmu DNA-sse, asendatakse see kohe tsütosiiniga. Tsütosiini asemel on metülatsioonist tingitud mutatsioonikiirus pisut suurenenud.
Samal ajal lülitatakse üha enam geene läbi metüleerimise, nii et rakuliinis olevad rakud muutuvad spetsialiseerunumaks. Remondiprotsessides põhinevad parandusensüümid algsel DNA ahelal, mida nad tunnevad ära kõrgema metüülimise kaudu. Ka täiendav tegevussuund on üles ehitatud sinna salvestatud teabe põhjal. Installimisvead parandatakse kohe. Lisaks katalüüsib ensüüm AID (aktiveerimisega indutseeritud tsütidiinaminaas) tsütidiinirühmade deaminatsiooni uridiinirühmadeks üheahelalises DNA-s. Toimuvad somaatilised hüpermutatsioonid, mis muudavad B-rakkude antikehajärjestusi. Seejärel valitakse sobivad B-rakud. See võimaldab paindlikku immuunvastust.
Haridus, esinemine, omadused ja optimaalsed väärtused
Tsütidiin on pürimidiini metabolismi vahesaadus. Kuna see on isoleeritud ühendus, pole see oluline. Nagu juba mainitud, koosneb see nukleiinalusest tsütosiinist ja neliku suhkru riboosist. Keha suudab ise tsütosiini sünteesida.
Selle süntees on aga väga energiamahukas, nii et see saadakse päästetee raames nukleiinhapete ehitusplokkidest ja seda saab uuesti integreerida nukleiinhapeteks. Kui alus on täielikult lagunenud, toodetakse süsinikdioksiidi, vett ja karbamiidi. See on RNA-s nukleosiidina. DNA-s on tsütosiin seotud desoksüboosiga, nii et nukleosiidi desoksütsütidiin toimub siin ehitusplokina.
Haigused ja häired
DNA tsütidiinijääkide metüülimine on märgistamisel väga oluline, et eraldada erinevad biokeemilised protsessid. Kuid vead võivad esineda ka metüleerimisel, mis põhjustavad haigust.
Defektiivsete metüülimiste korral võivad vallanduda nii suurenenud kui ka vähenenud geenitegevused, mis ei vasta nõuetele. Need metüülimismustrid antakse edasi raku jagunemise ajal. Pikemas perspektiivis toimuvad muutused, mis võivad põhjustada haigusi. Näiteks on mõnel kasvajarakul erinevad metüülimisstruktuurid, mida ei esine tervetel rakkudel. Näiteks võib metüleerimine blokeerida teatud geene, mis kodeerivad kasvu reguleerivaid ensüüme. Kui need ensüümid puuduvad, võib toimuda rakkude pidurdamatu kasv. See kehtib ka ensüümide kohta, mis põhjustavad raku normaalse surma (apoptoosi) raku defektide ilmnemisel.
DNA metüülimise sihipärane mõjutamine pole täna veel võimalik. Siiski on olemas uuringud kasvajarakkude täieliku demetüülimise kohta, et allutada need uuesti kasvu reguleerivate valkude kontrollile. Mitmete kliiniliste uuringute kohaselt on demetüleerimine tõestanud, et see piirab tuumori kasvu ägeda müeloidse leukeemiaga patsientidel. Seda protseduuri tuntakse ka kui epigeneetilist teraapiat. Metüleerimisprotsessid võivad mängida rolli ka teiste haiguste korral. Keskkonnamõjude tõttu kohaneb organism muutunud tingimustega bioloogiliste modifikatsioonide moodustumisega, mis põhinevad DNA tsütidiinijääkide metüülimisel. Keha viib seega läbi õppeprotsessi, mis võib aga põhjustada ka ebaõiget reguleerimist.
.jpg)