Adeniin "Heterobitsükliline aromaatne ühend, millel on puriini selgroog, mis orgaanilise tuuma alusena koos kolme teise alusega moodustab DNA-s ja RNA-s ühe geneetilise teabe põhiosa.
Lisaks mängib nukleosiidi või nukleotiidi kujul olev adeniin olulist rolli NAD, FADH2 või ATP metabolismis, eriti rakkude energiabilansis, mitokondrites.
Mis on adeniin
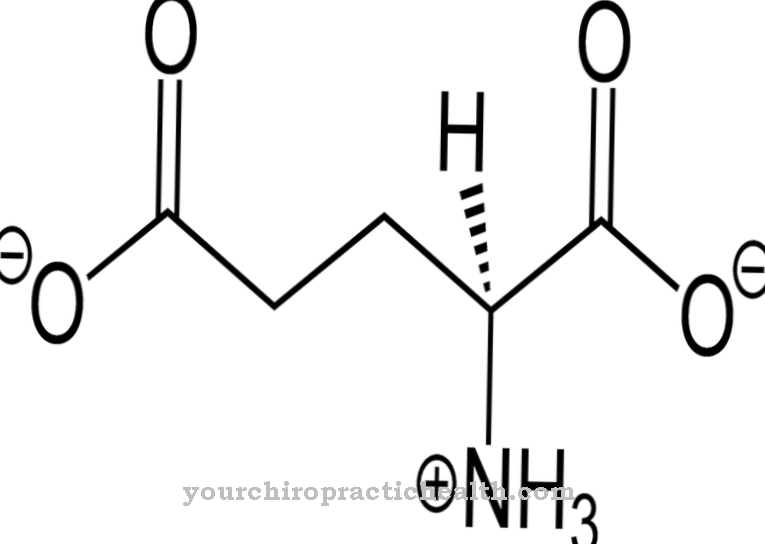
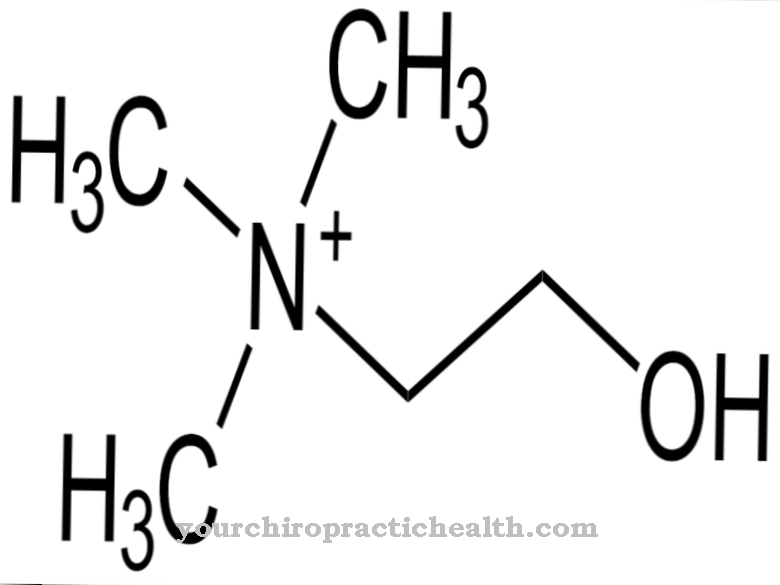
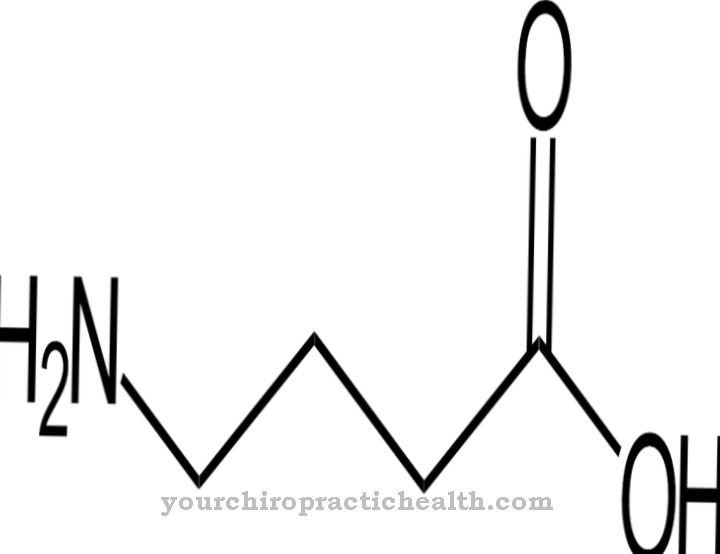
Adeniin keemilise molekulaarse valemiga C5N5H5 koosneb heterobitsüklilisest aromaatsest tuumast (puriini põhistruktuur) koos seotud aminorühmaga (NH2). Seetõttu nimetatakse ka adeniini Aminopuriin määratud. See on kahvatukollane tahke aine, mis sublimeerub temperatuuril 220 ° C, st muutub otse gaasilises olekus ja lahustub vees vaid halvasti.
Deoksüribioosisuhkru molekuli lisamine muudab adeniini deoksüadenosiiniks, mis on üks neljast ehitusplokist, mis moodustavad topeltheeliksi DNA. Tümidiinist ja sellega seotud desoksüboosi molekulist moodustunud desoksütümidiin toimib komplementaarse alusena. RNA puhul on see veidi muudetud protsess. Adeniinist saab D-riboosisuhkru molekuli lisamisega adenosiin. Adenosiin võtab RNA-s deoksüadenosiini positsiooni DNA-s. Komplementaarseks aluseks pole tümiin, vaid uratsiil uriini kujul.
Lisaks sellele moodustab adenosiin nukleotiidide ATP, ADP ja AMP põhistruktuuri, millel on oluline roll rakkude energiabilansis. Adenosiin täidab ka olulisi funktsioone kofaktorina paljudes ensüümides, hormoonides ja neuromodulaatorites nagu koensüüm A, NADPH ja NADH.
Funktsioon, mõju ja ülesanded
DNA topeltheeliksi ahela osana moodustab adenosiin kahe vesinikusilla kaudu adeniini-tümiini aluspaari (A-T) koos täiendava nukleiinaluse tümiiniga desoksütümidiini kujul. Enamasti üheahelalises RNA-s on adeniinil analoogne funktsioon, ehkki uratsiil ei ole tümidiin, kui komplementaarse aluse moodustamisel komplementaarseks aluseks olev mRNA (messenger RNA).
DNA ja RNA komponendina ei osale adeniin otseselt metaboolsetes protsessides, kuid seda kasutatakse koos teiste nukleobaasidega aminohapete järjestuste kodeerimiseks vastavate valkude sünteesiks. Peaaegu kõigi rakkude energiavahetuse osa, mida nimetatakse hingamisahelaks, koosneb sisuliselt oksüdatsiooni- ja redutseerimisprotsesside seeriast, nn redoksprotsessidest. Adenosiin, mis fosforüleeritakse adenosiintrifosfaadi (ATP) moodustamiseks, on hingamisahelas keskse tähtsusega. ATP vabastab fosforirühma ja saab seega adensoindifosfaadiks (ADP) või adenosiinmonofosfaadiks (AMP). Üldiselt on see eksotermiline protsess, mille käigus süsivesikute lagundamine toimub ainevahetuse energia tootmiseks ja nt. B. näeb ette lihaste tööd.
Selles funktsioonis osaleb adeniin või adenosiin otseselt keemilistes reaktsioonides. Redoksreaktsioonide ahela oluliseks dünaamiliseks komponendiks on ka elektronide ülekandmine vesinikuga (H) või muudele elektronkandjatele seotud elektronidest. Ka siin on adeniin ja adenosiin ensüümide või katalüsaatorite, näiteks nikotiidiamiidi (NAD) jt funktsionaalsed komponendid, mis lõpuks vesiniku vesiniku oksüdeerimise (põlemise) veeks jaotab paljudeks katalüütiliselt juhitavateks üksikuteks etappideks ja on seetõttu ainevahetusele kättesaadavad, põhjustamata põlemiskahjustusi. .
Haridus, esinemine, omadused ja optimaalsed väärtused
Keemilise valemi C5N5H5 kohaselt koosneb adeniin põhilistest ehitusplokkidest, süsinikust, lämmastikust ja vesinikust, mis kõik on looduses ohtrad. Haruldasi mikroelemente ega mineraale pole vaja. Vastavalt sellele ei pea kartma sünteesi toormaterjalide puudust, vaid pigem probleemi enda keha tootmisprotsessis.
Kuna süntees on aeganõudev ja energiamahukas, siis umbes 90% ajast kasutab keha teistsugust rada, sünteesib see adeniini ringlussevõtu teel. Puriini metabolismi käigus saadakse adeniin keerukamate ühendite laguproduktina. Adeniin muutub nukleosiidina biokeemiliselt efektiivseks ainult desoksüribose molekuli lisamisega. See muudab adeniini deoksüadenosiiniks. Ühe kuni kolme fosfaatjäägi täiendava lisamisega saab deoksüadenosiinist nukleotiid, mida nimetatakse adenosiinmonofosfaadiks (AMP), tsükliliseks adenosiinmonofosfaadiks (cAMP), adenosiini difosfaadiks (ADP) või adenosiintrifosfaadiks (ATP).
Vastavalt adeniini mitmekesistele ülesannetele ja selle biokeemiliselt aktiivsetele ilmingutele keskkonnas, mis vastab dünaamiliselt muutuvatele nõuetele ja vaba adeniini ei esine keha vereringes, ei saa adeniini taset mõõta. Puhta puriini metabolismi kohta saab kaudseid järeldusi teha ainult teatud metaboolseid protsesse jälgides ja mõõtes.
Haigused ja häired
Tuntuim - de facto, kuid haruldane - ainevahetushäire, mis ilmneb seoses keha enda toodetud adeniini ja selle bioaktiivsete vormidega, on Lesch-Nyhani sündroom. See on x-kromosoomi geneetiline defekt. Geenimutatsioon põhjustab hüpoksantiin-guaniini fosforibosüültransferaasi (HGPRT) täielikku defitsiiti.
HGPRT puudumine põhjustab puriini metabolismi häireid, mistõttu puriini aluste hüpoksantiini ja guaniini ringlussevõtt, mis tavaliselt toimub, pole enam vajalik. Selle asemel on keha sunnitud neosünteesi kaudu pidevalt adeniini tootma. See põhjustab liigset kusihappe kogust ja kusihappe kristallide sadestumist, mis võib põhjustada podagra või kusekivide moodustumist liigestes. Lisaks on vastsündinutel tavaliselt vaimse arengu defitsiit ja suurenenud autoagressioon.
Veel üks haruldane pärilik haigus on Huntingtoni tõbi. 4. kromosoomis on geneetiline defekt. Alusjärjestus tsütosiin-adeniin-guaniin 10 kuni 30 kordusega leitakse seal tavaliselt teatud geenis. Kui geenimutatsiooni tõttu on neid niinimetatud kolmikute kordusi rohkem kui 36, tekib pärilik haigus Huntingtoni tõbi. Haiguse käigus on motoorseid probleeme ja närvikahjustusi, mida ei saa ravida.








.jpg)
.jpg)